De minuscules grenouilles sans oreilles qui entendent fort bien
Certaines espèces de minuscules grenouilles ne possèdent ni orifice auditif, ni oreille moyenne, ni tympan, mais perçoivent cependant parfaitement les signaux acoustiques émis par leurs congénères. Une équipe de chercheurs de plusieurs laboratoires français dirigée par Renaud Boistel a pu, sur une espèce, Sechellophryne gardineri, vivant dans l’archipel des Seychelles, mesurer les propriétés biomécaniques des tissus auditifs de ces batraciens. Leur modélisation a pu montrer que c’est par excitation résonante de la cavité buccale que le son était perçu par l’oreille interne de ces animaux.

Fig.1 Photographie d’une grenouille S. gardineri . La barre d’échelle
mesure 5 mm. Crédit PNAS. R. Boistel.
Des batraciens sans oreille moyenne qui s’entendent très bien
Ces petites grenouilles mesurent environ 1,5 cm de long. On suppose que l’espèce a été isolée dans l’archipel des Seychelles, dans l’Océan Indien, il y a quelques 50 millions d’années, à la suite des fracturations du supercontinent nommé Gondwana à la fin de l’ère Jurassique.
Les sons émis par les grenouilles ont été enregistrés, puis rejoués dans leur habitat naturel. Cela a provoqué des vocalisations de la part de plusieurs grenouilles mâles. Ceci montre bien que ces batraciens sont capables d’entendre. En outre, les mâles ne répondaient pas à des appels d’autres espèces, ce qui indique de bonnes capacités de discernement acoustique. Il en est de même pour les Atelopus franciscus, espèce très voisine de S. Gardineri vivant en Amérique du Sud et que l’équipe avait déjà étudiée en Guyane française.
La recherche du trajet du son dans le corps de ces grenouilles
Quel pouvait donc être le chemin suivi par le son pour parvenir à l’oreille interne de ces animaux sans orifice auditif ni oreille moyenne? On avait proposé plusieurs pistes : la transmission par les poumons, celle par un osselet dit operculaire situé contre la peau à l’intérieur de la grenouille et enfin la conduction osseuse.
Pour répondre à cette question, il était bien difficile de faire des mesures acoustiques directes sur ces petits batraciens. On a donc cherché à obtenir une simulation numérique de la réponse à une excitation acoustique. des divers organes de la grenouille. Pour cela, il fallait connaître très précisément la masse spécifique en tout point de l’animal.
Pour cela, les chercheurs ont tout d’abord effectué une cartographie 3D complète de ces animaux et obtenu la connaissance exhaustive des masses spécifiques de tous les tissus biologiques les constituant. Sur un si petit animal, les techniques de tomographie X ou de scanner classiques n’avaient pas une assez grande résolution. Aussi, on a utilisé une technique de tomographie nouvelle, l’holotomographie, en faisant appel à une source de rayons X particulière, celle du synchrotron de l’ESRF (European Synchrotron Facility, Grenoble).
L’holotomographie a pour principale caractéristique d’être sensible, outre à l’amplitude du signal transmis à la phase de celui-ci. Ceci permet d’obtenir des images en contraste de phase. On perçoit alors des détails inaccessibles au mode en amplitude seul.
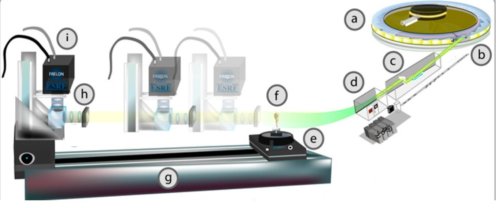
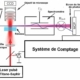
Fig.2. Schéma de l’appareillage expérimental utilisé pour la tomographie par rayonnement synchrotron.
a) Source de rayonnement synchrotron de l’ERSF Grenoble. Le faisceau de rayons X de forte intensité sort en b). En c) un monochromateur sélectionne des photons X d’une énergie proche de 20,5 keV(correspondant à une longueur d’onde de 60 picomètres, soit 60 millionièmes de micromètres). On obtient en d) un faisceau d’une section de 600 mm2 et d’une grande cohérence spatiale. e) Dispositif expérimental d’holotomographie. f) Le spécimen de grenouille peut tourner autour d’un axe vertical. g) distance qu’on peut faire varier entre le spécimen et le convertisseur h) de rayons X en lumière visible. i) Caméra CCD. Crédit PNAS. R. Boistel.
On voit sur la figure 3 des images de la tête de S. gardineri obtenues par holotomographie.
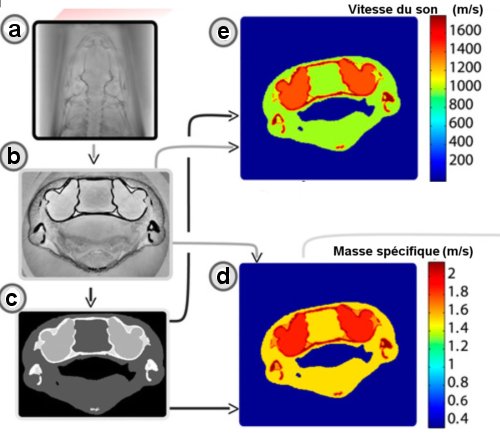
Fig.3. Quelques images 2D de la tête de S. gardineri obtenues par holotomographie.
a) Carte de phase obtenue à partir de 4 radiographies à
des distances différentes.
b)Tranche holotomographique utilisant les cartes de phase.
c) Segment d’image au niveau de l’oreille interne.
d) Carte de masse spécifique extraite des images holographiques.
e) Idem pour la vitesse du son. Crédit PNAS. Boistel.
Les images précédentes représentent des tranches à 2 dimensions fournies par la tomographie. La vidéo suivante montre une reconstruction 3D de l’oreille obtenue par les mêmes techniques sur une grenouille Atelopus franciscus.
[jwplayer mediaid= »14796″] L’oreille interne est figurée en vert. Crédit PLOS one. R.Boistel
Le trajet réel du son
En utilisant les cartes de masse spécifique et de vitesse de son obtenues sur Sechellophryne gardineri, les chercheurs ont simulé à l’ordinateur la propagation du son à travers la tête de l’animal. S. gardineri produit un signal sonore d’avertissement vers 5700 Hz, c’est donc autour de ces fréquences qu’ont été réalisées les simulations.
L’analyse holotomographique montre que le système operculaire de cette espèce est bien développé, mais que sa réponse acoustique est limitée à des fréquences inférieures à 1000 Hz.
On peut aussi écarter un système d’ouïe basé sur les poumons, car l’analyse montre que les fréquences de résonance correspondantes sont du domaine des ultrasons et donc incompatibles avec les fréquences émises et perçues par cette espèce.
La conduction osseuse pouvait constituer une alternative dans de si petits animaux, mais elle apparût insuffisante pour expliquer le transport du son vers l’oreille interne.
Et c’est la cavité orale qui joue le rôle principal. En effet, sa fréquence de résonance de 5738 Hz est très proche de celle de l’appel de S. gardineri. La bouche amplifie le signal sonore par résonance et on remarque en outre que les tissus séparant la cavité buccale de l’oreille interne sont d’une finesse extrême (80 micromètres) au contraire de ce qui existe dans les espèces de grenouilles qui possèdent une oreille externe tympanique. On peut dire pour résumer que ces animaux entendent par la bouche!
En théorie de l’évolution, on pensait jusqu’ici que le passage pour les tétrapodes (vertébrés possédant deux paires de membres) d’un mode de vie aquatique à un mode terrestre s’était toujours accompagné d’une évolution de l’oreille moyenne vers une oreille tympanique, solution la plus polyvalente pour la vie terrestre. Ces résultats montrent qu’il n’en est rien. On ne peut qu’admirer le caractère multidisciplinaire de cette étude qui a réuni biologistes, physiciens, paléontologues et ingénieurs. Voici en effet la liste des laboratoires auxquels sont affiliés les chercheurs de l’étude:
Centre de Neurosciences Paris-Sud (CNPS), CNRS UMR 8195, Université Paris XI,Orsay ;
Université Paris Sud, Orsay;
Institut International de Paléoprimatologie et de Paléontologie Humaine (IPHEP), CNRS UMR7262, Université de Poitiers, Poitiers;
European Synchrotron Radiation Facility, 38043 Grenoble, France;
Centre de Recherche en Acquisition
et Traitement de l’Image pour la Santé, CNRS UMR 5515, Institut national des sciences appliquées de Lyon, 69621 Villeurbanne, France;
Laboratoire de Mécanique et d’Acoustique (LMA), CNRS UPR 7051, 13402 Marseille, France;
Institut de Biologie Systémique et Synthétique (iSSB), Genopole, CNRS, Université d’Evry Val d’Essonne, Evry;
Institut Langevin, CNRS UMR 7587, ESPCI de la ville de Paris ParisTech, Paris.
Pour en savoir plus :
How minute sooglossid frogs hear without
a middle ear
Renaud Boistel, Thierry Aubin, Peter Cloetens, Françoise Peyrin, Thierry Scotti, Philippe Herzog,
Justin Gerlach, Nicolas Pollet, and Jean-François Aubry PNAS,septembre 2013
Whispering to the Deaf: Communication by a Frog without External Vocal Sac or Tympanum in Noisy Environments
Renaud Boistel, Thierry Aubin, Peter Cloetens, Max Langer, Brigitte Gillet, Patrice Josset, Nicolas Pollet, Anthony Herrel
PLOS One, juillet 2011, accès libre.